В статье обсуждается возможный способ разграничения симпатрических и аллопатрических популяций ряпушки нами был использован показатель отношения максимального количества счетных признаков (жаберных тычинок на первой жаберной дуге; чешуй в боковой линии) к минимальному числу для ряда озер России, Швеции, США, Канады.
Наряду с сигом Coregonus lavaretus complex, европейской ряпушке С. albula L. характерен исключительно сложный морфологический и экологический полиморфизм. Как известно относительно видового статуса С. albula L. Евразии существуют, в основном две точки зрения. Часть исследователей (Решетников, 1980) признает, что в Евразии существует один полиморфный вид ряпушки С. albula L. По мнению других исследователей, точку зрения которых мы разделяем (Китаев, 1993), вид С. albula L. представлен несколькими видами.
Подобно работе проведенной С. П. Китаевым для вида Coregonus lavaretus L. и для гольца (Китаев, 2004), как способ разграничения симпатрических и аллопатрических популяций ряпушки нами был использован показатель отношения максимального количества счетных признаков (жаберных тычинок на первой жаберной дуге; количество чешуй в боковой линии) к минимальному числу. В результате на основании данных таблиц 1 и 2 озера были разбиты на три основные группировки: в водоеме один вид (форма) ряпушки – аллопатрия; вторая две и более формы; третьей группе озер два и более вида с формами – как проявление симпатрия.
Таблица 1. Отношение максимального числа жаберных тычинок к минимальному числу у аллопатрически и симпарически обитающих ряпушек (Беляева, 1953; Берг, 1949; Лукаш, 1939; Дрягин, 1949; Скрябин, 1977; Лопатышкина, 1957; Scott., Crossman, 1973; Дятлов, 2002).
|
Аллопатрические популяции |
Симпатрические популяции одного вида |
Симпатрические формы разных видов
|
|||||
| Европейская ряпушка | Сибирская ряпушка | ||||||
| Водоем | Max/min
(колеб) |
Водоем | Max/min
(колеб) |
Водоем | Max/min
(колеб) |
Водоем | Max/min
(колеб) |
| Оз. Сиклоя (Швеция) | 1,23 (39-48) | Р. Колыма (Россия) | 1,23 (38-47) | Ладожское озеро (Россия)) | 1,48 (37-55) | Эри (США) | 1,76 (30-53) |
| Оз. Халсьён (Швеция) | 1,07 (42-45) | Р. Лена (Россия) | 1,36 (38-52) | Онежское озеро (Россия) | 1,26 (38-48) | Гурон (США) | 2,12 (25-53) |
| Оз. Асунден (Швеция) | 1,02 (43-44) | Р. Норильская (Россия) | 1,37 (37-51) | Оз. Шарташ (Россия) | 1,51 (35-53) | Мичиган (США) | 2,00 (26-52) |
| Оз. Феген (Швеция) | 1,07 (42-45) | Р.Пясина | 1,54 (37-57) | Онтарио (США) | 1,63 (33-54) | ||
| Оз. Орен (Швеция) | 1,02 (43-44) | Р. Лама | 1,30 (36-47) | Верхнее (США) | 1,65 (32-53) | ||
| Оз. Аснен (Швеция) | 1,21 (38-46) | Р. Щучья (Россия) | 1,20 (34-41) | Нипигон (США) | 1,50
(32-48) |
||
| Оз. Августовское (Россия) | 1,10
(42-47) |
Р. Лена (Россия) | 1,36 (38-52) | Оз. Виннипенг (Канада) | 1,31 (41-54) | ||
| Беломор.ряпушка (Россия) | 1,13 (36-41) | Р. Колыма (Россия) | 1,37 (35-48) | Оз. Атлин (Канада) | 1,26 (42-53) | ||
| Оз. Керетьозеро (Россия) | 1,28 (35-45) | Р. Анадырь (Россия) | 1,25 (40-50) | Оз. шт. Аляска (США) | 1,26 (42-53) | ||
| Топозеро (басс. Белое море) (Россия) | 1,21 (41-50) | Р. Амгуэма (Россия) | 1,30 (43-56) | Медвежье озеро (Канада) | 1,21 (41-50) | ||
| Урос (Россия) | 1,26 (38-48) | Р. Аччен (Россия) | 1,10 (45-50) | ||||
| Вендерское (Россия) | 1,08 (34-37) | Р. Иони (Россия) | 1,04 (44-46) | ||||
| Риндозеро (Россия) | 1,07 (40-43) | Оз. Баунт (Россия) | 1,20 (40-48) | ||||
| Вахваярви (Россия) | 1,30
(35-46) |
Оз. Арылых | 1,26 (38-48) | ||||
| Валкеалампи (Россия) | 1,32 (34-45) | Оз. Первое Пуринское | 1,31 (35-46) | ||||
| Топозеро(басс.Лад.оз)(Россия) | 1,18 (33-49) | Оз. Собачье (Россия) | 1,23 (39-48) | ||||
| Куйкаярви (Россия) | 1,25 (36-45) | Оз. Кета (Россия) | 1,37 (37-51) | ||||
| Чужмозеро (Россия) | 1,36 (36-49) | Оз. Таммот | 1,24 (41-51) | ||||
| Оз. Люцин (Россия) | 1,20
(39-47) |
Обская губа (Россия) | 1,20 (34-41) | ||||
| Водлозеро (Россия) | 1,17 (35-41) | Тазовская губа (Россия) | 1,20 (34-41) | ||||
| Переславское озеро (Россия) | 1,28 (42-54) | ||||||
Таблица 2. Отношение максимального числа чешуй в боковой линии к минимальному числу у аллопатрически и симпарически обитающих ряпушек (Беляева, 1953; Дятлов, 1983, 1993; Беляева, 1953; Берг, 1949; Лукаш, 1939; Дрягин, 1949; Скрябин, 1977; Scott, Crossman,1973).
| Европейская ряпушка | Сибирская ряпушка | Симпатрические популяции одного вида | Симпатрические формы разных видов | ||||
| Водоем | Max/min
(колеб) |
Водоем | Max/min
(колеб) |
Водоем | Max/min
(колеб) |
Водоем | Max/min
(колеб) |
| Урос (Россия) | 1,30
(68-91) |
Р. Пясина (Россия) | 1,44
(77-111) |
Ладожское озеро (Россия) | 1,40
(62-89) |
Эри
(США) |
1,31
(63-83) |
| Вендерское (Россия) | 1,06
(77-82) |
Р. Лена (Россия) | 1,20
(79-95) |
Онежское озеро (Россия) | 1,40
(63-91) |
Гурон (США) | 1,49
(63-94) |
| Риндозеро (Россия) | 1,05
(75-79) |
Р. Колыма (Россия) | 1,32
(75-99) |
Оз. Увильды (Россия) | 1,34
(67-90) |
Мичиган (США) | 1,41
(63-89) |
| Вахваярви (Россия) | 1,26
(76-96) |
Р. Енисей (Россия) | 1,20
(69-83) |
Оз. Шарташ (Россия) | 1,47
(73-108) |
Онтарио (США) | 1,49
(63-94) |
| Валкеалампи (Россия) | 1,18
(75-89) |
Р. Амгуэма (Россия) | 1,25
(80-100) |
Верхнее (США) | 1,49
(63-94) |
||
| Топозеро (басс. Ладож. озера) | 1,38
(62-86) |
Р. Анадырь (Россия) | 1,27
(73-93) |
Нипигон (США) | 1,29
(64-83) |
||
| Куйкаярви (Россия) | 1,31
(73-96) |
Оз. Лама (Россия) | 1,22
(73-89) |
Винипег (Канада) | 1,18
(58-69) |
||
| Чужмозеро (Россия) | 1,31
(70-92) |
Оз. Собачье (Россия) | 1,24
(77-96) |
Атабаска (Канада) | 1,18
(58-69) |
||
| Водлозеро (Россия) | 1,22
(70-86) |
Оз. Баунт (Россия) | 1,33
(72-96) |
||||
| Переславское озеро (Россия) | 1,17
(76-89) |
Оз. Арылах (Россия) | 1,28
(70-90) |
||||
| Оз. Первое Пуринское (Россия) | 1,40
(74-104) |
||||||
| Оз. Таммот (Россия) | 1,31
(67-88) |
||||||
В сравнении участвовали три ареала ряпушек: 1. Сибирские ряпушки – как предполагаемый центр происхождения всех сиговых; и два основных ответвления (с разными путями дальнейшей эволюции): 2. Ряпушки Европы; 3. Североамериканские ряпушки. В итоге мы получили шесть объединений: 1. Аллопатрические популяции ряпушки (Coregonus albula L.) – некрупные озера; 2. Симпатрические популяции ряпушки (Coregonus albula L. + Coregonus albula (infraspecies) kiletz Michailowsky) для Онежского озера; Coregonus albula L + Coregonus albula (infraspecies) ladogenis Pravdin) для Ладожское озера; 3. Сибирская ряпушка (речная) (Coregonus sardinella Valenciennеs); 4. Сибирская ряпушка (озерная) (Coregonus sardinella Valenciennеs). 5. Озера Канады и штата Аляска (США); 6.Великие Американские озера (США) (рис. 1).
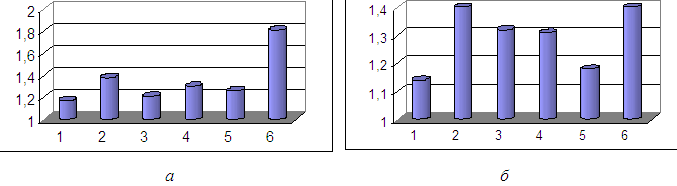
Рисунок 1. Сравнительная характеристика отношения максимального числа к минимальному а-жаберных тычинок; б – числа чешуй в боковой линии с целью нахождения количества сосуществующих видов в одном водоеме
Как следует из рисунка 1-а аллопатрические популяции ряпушки имеют значительно более низкий показатель max-min в отличие от симпатрических. Сибирские ряпушки имеют достаточно высокий показатель max-min. У ряпушек Великих американских озер данный показатель здесь наибольший среди всех групп. А, как известно данные озера в своем составе имеют несколько видов ряпушек. Другой пример, ряпушка оз. Шарташ имеет кроме ранее акклиматизированного рипуса ещё и гибрид рипуса с чудским сигом, что возможно нашло свое отражение в max-min показателе (табл. 1). Кроме того, согласно рисунку, североамериканская ряпушка, по продвижению с юга на север от Великих американских озер показатель max-min снижается, и становиться более схож с ряпушками Евразии. Из рисунка 1-б видно, что тенденция характерная для первого случая (рис. 1-а) также хорошо прослеживается. На основании этого данный показатель также можно рекомендовать для оценки количества сосуществующих видов в одном водоеме. При этом подсчет количества чешуй в боковой линии является более простым, чем подсчет жаберных тычинок, хотя для полной картины надо просчитывать и их.
Конечно же, в любом правиле есть свои исключения. В нашем случае это ряпушка оз. Чужмозеро, оз. Вахваярви, оз. Валкеалампи и Куйккаярви имевшие отношение соответственно 1,36, 1,30, 1,32 и 1,28. Не исключено, что ряпушка этих озер в процессе видообразования образовала две или более форм. Прежде всего, это относится к ряпушкам оз. Чужмозеро, оз. Вахваярви, т.к., в отличие от остальных двух, они имеют большую схожесть с европейской. Пусть и с меньшими, но довольно близкими значениями можно выделить ряпушку оз. Урос, оз. Переславского и оз. Керетьозеро. Возможно, они находятся на разных стадиях процесса аллопатрического видообразования. На наш взгляд, вся микроэволюция ряпушек идет по пути к образованию новых форм посредствам: увеличения ареала обитания и спектра питания (на более энергоемкий).
Для ряпушки оз. Куйккаярви Дятлов (1993) уже предложил выделять в две её формы: крупноглазую и мелкоглазую. Из 45 сравниваемых морфологических признаков данные формы достоверно различаются по 10. Коэффициент различия и процент неперекрывания частично перекрывающихся кривых у крупно – и мелкоглазых особей выявлен в массе тела, в длине по Смиту, длине без хвостового плавника и длине анального плавника. По четырем параметрам он выше принятого показателя подвидового различия. Кроме всего прочего, оз. Валкеалампи и Куйккаярви относятся к одной системе р. Ууксионъоки. Ещё одной причиной столь резкого отклонения от типичной формы европейской ряпушки для этих популяций можно объяснить следующим образом. Для ряпушки оз. Валкеалампи характерны признаки, сближающие их не с типичной европейской ряпушкой, а с сибирской (беломорской) ряпушкой по некоторым признакам. Антедорсальное расстояние не на много превышает 42% (до 42% – признак сибирской ряпушки), антевентральное расстояние 43- 44 % (до 45 % признак сибирской ряпушки), отсюда смещение плавников в передний отдел тела; высота спинного плавника 17-18%, особенно для оз. Куйккаярви (18,72+0,56) (15 % и более признак сибирской ряпушки) длина основания анального плавника 12-13 % (12% более признак сибирской ряпушки), высота анального плавника 11-12% (10% и более признак сибирской ряпушки) (Покровский, 1967).
Естественно, данное предположение требует дальнейшей проработки. Ряпушка Валкеалампи, скорее всего, берёт свое начало от ряпушки оз. Куйккаярви, между этими двумя озерами до сих пор существует связь (ручей длиной 250 м, и глубиной 20 см), но миграция по нему рыбы сейчас не представляется возможным (Дятлов, 1983, 1993). Очевидно, эта связь существовала (или периодически возникает в период наибольшего уровня воды). Взаимопереходы мелкой ряпушки в крупную хорошо известны (Потапова, 1978). Систематическое положение ряпушки Валкеалампи требует уточнения, все остальные исследованные популяции принадлежат к виду: европейская ряпушка Coregonus albula L. В водоемах Сибири достаточно часто встречаются популяции ряпушки более схожие с европейской ряпушкой.
Материалы О.Л. Ольшанской (1967), Ф.Н. Кириллова (1972) свидетельствуют о том, что переход речных ряпушек к жизни в озерах сопровождается в среднем увеличением антедорсального расстояния и уменьшением вентроанального. Отношение вентроанального расстояния к антедорсальному у некоторых популяция сибирской ряпушки еще остается средним между речными и озерными формами. Норильская ряпушка, и особенно кетинская, в целом отличаются от исходной пясинской ряпушки, и сближается с европейской ряпушкой сильнее, чем с сибирской. Это, по нашему мнению, связано с относительной молодостью обособления последней от речной и близостью их ареалов. На основании этого она приобрела морфологические признаки более похожими с европейской ряпушкой.
Поскольку главные диагностические признаки имеют сильную размерно-возрастную изменчивость у речных и озерных ряпушек, и поскольку озерные ряпушки европейской части России и Сибири меньше речных форм, то отмеченные изменения морфологии не только результат приспособления к новым условиям среды, но и результат корреляции с размерами тела.
Таким образом, переход сибирских ряпушек от проходного образа жизни к озерному и уменьшение размеров их тела в среде с меньшей динамичностью водной массы в конечном итоге привела к сближению в морфологии озерных сибирских ряпушек с европейскими. Следовательно, мы должны отметить, что признаки, на основании которых идет разделение подвидов ряпушки недостаточно устойчивые. В озерах с длительной историей существования независимо от их расположения возникают озерные более мелкие формы, близкие по морфологии европейским, а в действительности – популяциям ряпушек с озерным экотипом.
Не имея возможность рассмотреть здесь большое количество примеров и теоретические аспекты формирования подобных пучков видов у ряпушки, отметим, что с нашей точки зрения возможен вопрос – только ли в озерах имеются условия для формирования пучков симпатрических видов (форм) рыб или в определенных условиях их возникновение возможно и в водоемах других типов? К примеру, по данным Голубцова (2003) формирование пучков симпатрических видов или форм может происходить, как в озерных, так и в речных условиях. Промежуточные формы связаны с молодостью популяций или с гибридизацией озерных и речных форм в зоне стыка ареалов – беломорская ряпушка и ее популяции в отдельных водоемах европейской части России. Это свидетельствует о молодости вида Coregonus albula L., находящегося в стадии интенсивного формообразования и деления на ряд резко различных популяций, которые в дальнейшем могут дать начало новым систематическим категориям.
Поэтому необходимо найти более показательные характеристики, отделяющие сибирскую и европейскую ряпушку. Иначе ограничиться лишь констатацией того, что на всей территории России встречается как сибирская, так и европейская ряпушка, единственное, что на европейской части России больше встречается европейская ряпушка Coregonus albula L., а в ее азиатской части – сибирская Coregonus sardinella Valenciennеs.
Наиболее вероятной причиной дифференциации ряпушки на отдельные популяционные группировки, на наш взгляд, является влияние факторов окружающей среды. Они лежат в основе симпатрической дивергенции группы американских ряпушек (“cisco”) в Великих озерах (Todd, Smith, 1992). На процессы образования влияют температура, освещенность, давление, вязкость воды содержание кислорода, структура субстрата, сезонные изменения всех показателей, коррелирующие с глубиной водоема, характером питания и воздействием хищников. Комбинация этих факторов создает ступенчатость экологических градиентов. Дополняет это географическая структура самих озер постоянно меняющейся сетью заливов и прибрежной зоной, разделенной на участки с различными условиями, способствующими специализации в жизненном цикле, физиологии, поведении и морфологии. Ограниченность ресурсов в пространстве и времени, их разрывы приводят к возникновению демов, занимающих в результате сильного отбора различные адаптивные пики. Популяции, которые попадают между пиками, элиминируются отбором. По мнению Тодда и Смита (Todd, Smith, 1992), такой сценарий неоднократно повторялся в Великих озерах, в результате чего здесь возник целый букет репродуктивно изолированных видов или полувидов ряпушек, причем это происходило в сравнительно недавнее время.
Можно предположить, что сходные факторы оказывают влияние на формирование внутривидовой популяционной структуры сибирской ряпушки. Однако, по сравнению с Великими озерами, условия здесь более суровы, среднегодовая температура ниже, вегетационный период намного короче, кормовая база скуднее, число ниш меньше. Таким образом, в водоемах Сибири вид сибирская ряпушка представляет систему локальных, в той или иной степени изолированных, группировок со специфическими морфоэкологическими особенностями. Однако изолирующие механизмы под воздействием ряда причин (климатические, гидрологические факторы, техногенное воздействие, промысел, резкие природные колебания численности самой ряпушки, наличие проходной формы и.т.д.) непрерывно нарушаются. Это приводит к появлению “заболачивающего эффекта” (“swamping effect”) скрещивания между разными группировками и к возможности быстрого изменения фенотипа по внешним морфологическим признакам. В результате создается своеобразная буферная система, в которой процессы дивергенции подавлены. Ее динамичная структура, будучи крайне разнообразной, в целом хорошо адаптирована к условиям высоких широт.
На основании всего вышесказанного можно заключить, что в результате данных анализов мы могли видеть примеры симпатрического (Онежское и Ладожское озера) и аллопатрического (Урос, Чужмозеро) видообразования. Наши утверждения основываются на степени возможной панмиксии форм ряпушки, показатель которой в первом случае гораздо менее вероятен; дифференциация по биологическим и части морфологических показателей существует.
Следовательно, симпатрическое видообразование в северных районах может иметь место лишь при условии, если водоем (озеро) достаточно велик и, на основании этого, имеет различные в ареале, условиях обитания, особенностями жизненного цикла (глубины, гидрохимия и.т.п.). Относительно больших показателей min-maх у сибирских ряпушек, то на наш взгляд этот связано с тем, что реки Сибири берут свое начало с южных районов степень формообразования, где значительно выше, чем в северных районах (Дгебуадзе, 2001). На этом, возможно, основывается, столь бурное видообразование в Великих озерах США. Подтверждением этому служит, при продвижении на север от Великих Американских озер, оз. Барроу (Канада) показатель max-min, где не превышает 1,36, оз. Виннипег (Канада) 1,31. оз. Атлин (Канада) и озера штата Аляска (США) 1,26; Большое Медвежье озеро (Канада) 1,21. В лучшем случае, здесь обитают лишь 1-2 формы ряпушки.
На основании всего вышесказанного, можно заключить, что в Онежском и Ладожском озерах обитают 2 вида, или 1 вид и 1 группа завершающая процесс видообразования. Относительно сибирских ряпушек, вопрос и наличии здесь нескольких видов (какого- либо видообразования) является вопросом для последующих изучений, хотя на наш взгляд, по крайней мере, ранг биологической формы, а в некоторых случаях и биорасы, им можно причислить. Таким образом, для разграничения симпатрических и аллопатрических популяций ряпушки лучше всего основываться на отношении максимального числа жаберных тычинок к его минимальному числу.
Библиографический список
- Беляева К.И. Экологические и биологические особенности крупных ряпушек из озер Карело-Финской ССР // Автореф. дис. на соиск. уч. ст. канд. биол. наук. Петрозаводск. 1953. 16 с.
- Берг Л.С. Рыбы пресных вод СССР и сопредельных стран // М., Л.: Изд-во АН СССР. Ч. 1 1949. С. 470 – 925.
- Голубцов А.С. Рыбы Эфиопии: распространение, систематика, эволюция // Дисс. на соиск. науч. степени доктора биологических наук. М. 2003. С 58.
- Дгебуадзе Ю.Ю. Экологические закономерности изменчивости роста рыб. М.:Наука, 2001., 274 с
- Дрягин П.А. Ряпушка Coregonus albula Linne // Промысловые рыбы СССР. М. 1949. С.218-222.
- Дятлов М.А. Ряпушка оз. Вахваярви // Лососевые Карелии. Петрозаводск. 1983. С. 117-126.
- Дятлов М.А. Ряпушка Coregonus albula озер системы р. Уксионьеки Ладожского озера // Проблемы лососевых на Европейском севере. Петрозаводск. 1993. С. 57-74.
- Дятлов М.А. Рыбы Ладожского озера. - Петрозаводск. 2002. 280 с.
- Кириллов Ф.Н. Рыбы Якутии. М.: Наука, 1972. 360с.
- Китаев С.П. К вопросу о систематике и распространении сиговых (Coregonidae) // Проблема Лососевых на Европейского Севере. 1993. С. 4-35.
- Китаев С.П. О сходстве морфологии, экологии, кариологии и явление парелеллизма, дивергенции и конвергенции сиговых и гольцов // Препринт. 2004. 47 с.
- Лопышкина Г.М. Морфологическое изменение рипуса при его акклиматизации в озерах Урала // Тр. ГосНИИ озерн. и речн. хоз-ва. 1957 т. 34. С 60 – 64.
- Лукаш Б. Рыбное хозяйство Карелии. Том V, 1939. 130 с.
- Ольшанская О.Л. Ряпушка бассейна реки Пясины / Тр. Красноярск. отд. СибНИИРХ, 1967. Т.9. С.94-213
- Покровский В. В. О морфологических особенностях, происхождении и географическом распространении беломорской ряпушки Coregonus sardinella maris-albi Berg // Изв. ГосНИОРХ. 1967. Т. 62. С. 100–114.
- Потапова О.И. Крупная ряпушка Coregonus albula (L) // под ред. Ю.А. Смирнова. Л. Наука. 1978. 113 c.
- Решетников Ю.С. Экология и систематика сиговых рыб. М.,: Наука. 1980. 300 с.
- Скрябин А.Г. Рыбы Баунтовских озер Забайкалья. М.:Наука, 1977, 230 с.
- Scott W.B., Crossman E.J. Freshwaters fishes of Canada. - Bull. Fish. Res. Canada, 1973 № 184, 996 p.
- Todd T.N., Smith G.R. A review of differentiation in Great Lakes Ciscoes // Pol. Arch. Hydrobiology. 1992. Vol. 39, № 3/4. Р. 261-267.