Как у всех организмов, находящихся в процессе видообразования, решающие события развертываются, вероятно, в изолированных, периферических популяциях, которые не были достаточно изучены. Фенотипические изменения популяций отмечались многократно, в частности, при изменении гидрологического режима водоемов и при переселении группы особей в водоем, где ранее представители данного вида не обитали. Анализ литературных данных показывает, что фенетическим следствием изменения условий обитания оказываются изменения темпа роста особей, их предельные размеры и определяемые различия и темпе роста изменения пластических признаков (Никольский, 1980; Яблоков, 1981). Как отмечал М.В. Мина (1986), “возникновение внутрипопуляционных различий в характере роста и развития особей представляется весьма вероятной предпосылкой симпатричекого видообразования уже потому, что влечет за собой возникновение различий по множеству эпигенетически связанных признаков, среди которых могут оказаться определяющие ассортативность и отбор.
Рассмотрим предполагаемую генетическую историю новообразования популяции, пространственно изолированной от той родительской популяции в условиях крупного водоема (Онежское и Ладожское озера), от которой она ответвилась. В качестве рабочей гипотезы можно допустить, что для такой изолированной популяции новый ареал может стать основным, оказавшись более благоприятным в измененной экологической ситуации на краю ареала. При этом у видов, приспособившихся к обитанию в одном ареале, степень наследственной изменчивости низка. Предположим, что они имеют одинаковые потребности в условиях существования и новая популяция вначале полностью генетически идентична родительской популяции, хотя, как известно, основатели новой популяции неизбежно содержат лишь малую часть общей изменчивости родительского вида (Mayr, 1982; Бердников, 1990).
Новые местные условия создают новые давления отбора, усиливающие непрерывное изменение генного состава и ведущие к развитию многочисленных новых приспособлений. Чем меньше размер популяции, тем, видимо, более она подвержена опасности вымирания. (Бердников, 1990). Следовательно, первым естественным свойством новой популяции является адаптация и увеличение ареала. Величина фенотипических различий между таксонами оказывается зависящей от длительности периода интенсивных фенотипических изменений. Конечно же, нельзя говорить, что этот процесс обязательно пойдет по данному пути т.к. видообразование процесс медленный, исторический, и ни один исследователь не может наблюдать его непосредственно (исключение случаи полиплоидии), а может наблюдать лишь в определенные фиксированные фазы, т.е. мы созерцаем лишь моменты в “преобразованиях видов”. Возможно, при изменении каких-нибудь условий, процесс дальнейшего видообразования приостановиться или популяция примкнет к родительской популяции, т.е. процесс видообразования непрерывен, но его направление зависит от условий, которые приобретет или не приобретет та или иная экосистема. Если в результате разрушения изолирующих преград бывший изолят восстанавливает контакт с родительским видом до того, как закончилось формирование изолирующих механизмов, в области контакта между этими формами образуются гибридная зона. При этом приобретение за период изоляции различия в биотопических требованиях могут уменьшить ширину гибридной зоны. Вторжение вновь образовавшегося вида в область распространения родительского или сестринского вида служат решающим доказательством того, что этот новый вид закончил свой процесс видообразования (Майр, 1974). Популяция основателей отличается от родительской популяции не только крайне малым разнообразием генофонда, но также и тем, что она подвержена действию совершенно нового сочетания факторов среды, как биотических, так и генетических. Новая изолированная популяция, возникшая вследствие изоляции, вполне может принять характер цепной реакции. Однако реакция этой популяции на давление отбора будет совершенно иной, чем реакция популяции, представляющей собой одну из непрерывного ряда соприкасающихся друг с другом популяций, связанных потоком генов и всеми интегрирующими механизмами. Чем дольше популяции находились в изолированном состоянии, тем в более резкой форме протекает генетическая революция, и чем больше экологические различия между областями, тем больше вероятность возникновения экологических различий между популяциями (Шмидт-Ниельсен, 1987; Бердников, 1990). Отбор в природе даже в самой малой популяции направлен, прежде всего, к общей приспособленности. Со временем такая изолированная популяция может не только полностью адаптироваться к новому месту обитания, но в качестве побочного продукта географической изоляции и генетических изменений в популяции приобрести и репродуктивную изоляцию от родительского вида. Таким образом, специализация к определенному ареалу создает идеальные предпосылки для быстрого видообразования, т.е. возникает адаптационный полиморфизм (Яблоков, 1981).
В дальнейшем эти, как минимум, две популяции, как только они разделятся, по ряду причин начнут расходиться и по генетическому составу, и различия между ними будут увеличиваться. Так4им образом, расширение ареала может повлечь за собой возрастание давления отбора, что, в свою очередь приведет к быстрой генетической перестройке популяции. Вероятность того, что одни и те же мутации в одинаковой последовательности будут происходить в всех популяциях, равна нулю. Каждая мутация, включенная в генотип, изменяет генетический фон популяции и тем самым влияет на селективную ценность всех последующих мутаций. Кроме того, в результате рекомбинаций в этих двух генофондах возникнут разные генотипы и, следовательно, поскольку один и тот же ген может иметь различную селективную ценность в разных генотипах, произойдет постепенное изменение частот генов. Наконец, “генетическая неопределенность” приводит к дивергенции генотипов. Если несколько комбинаций генов имеют равную селективную ценность (по отношению к данному давлению отбора), то чистая случайность или из этих комбинаций в состав данного генофонда. А, как известно, любая дивергенция двух генофондов увеличивает различие в генетическом фоне всех генов этих двух популяций, и, следовательно, создаются новые давления отбора.
При этом необходимо помнить, что перестройка генного комплекса популяции может происходить только в том случае, если популяция, хотя бы временно защищена от притока чуждых генов. Конечно же, это легче всего достигается при помощи внешних преград, а именно через пространственную изоляцию. По-видимому, подобна пространственная изоляция, всегда поддерживается географическими преградами. Как только это достигнуто, вновь образовавшийся вид может вторгнуться в пределы ареала родительского вида и обитать рядом с ним. Следовательно, основным изолирующим механизмом, на первых этапах видообразования, являются местоположение, глубины, что в свою очередь ведет к различиям в гидрохимии, т.к. популяция обитает уже отдельно.
В данной связи одним из интереснейших представителями сиговых рыб Карелии являются ряпушки. У вида – европейской ряпушки Онежского и Ладожского озер выделяют, наряду с мелкой, ее крупные экологических формы кильца (Coregonus albula kiletz Michajlowsky) Онежского озера и рипуса (Coregonus albula ladogenus Pravdin) Ладожского озера, отличающиеся между собой по ряду биологических показателей (размерно-возрастная структура, плодовитость, сроки наступления половозрелости, типа питания) (Георгиев, 2004). Подобные популяции называются симпатрическими.
Для определения степени сходства и различия между симпатрическими популяциями Онежского и Ладожского озерах, на современном этапе, нами был проведен дискриминантный анализ кильца, ряпушки Онежского озера и рипуса, ряпушки Ладожского озера.
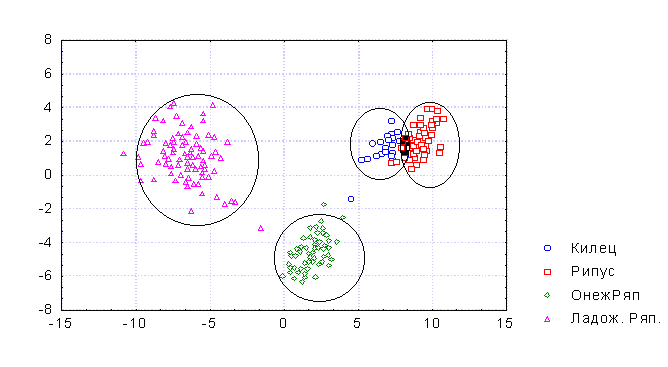
Рисунок 1. Характеристика различных популяций (форм) ряпушки Ладожского и Онежского озер на основе дискриминантного анализа.
Из рисунка (рис. 1) видно, что килец и рипус занимают соседствующее положение не со своими типичными мелкими формами, а по отношению друг к другу. Возможной причиной этого, на наш взгляд является то, что крупные формы являются неким завершающим звеном видообразования у ряпушек, и этот путь преобразования шел параллельно в Онежском и Ладожском озере. Следовательно, подобный результат микроэволюции ряпушек следует признать неслучайным и, как результат единого процесса, их сходство в морфологических признаках. Возможно, в дальнейшем будет происходить процесс дивергенции уже и кильца и рипуса. К примеру, Дятлов (2000) приводит пример того, что в Ладожском озере существуют уже две формы рипуса отличающиеся по ряду морфологических и биологических признаков. На наш взгляд, пока ранг биологической формы этим популяциям давать рано, т. к. их основные отличия лежат в различных ареалах (северный – глубоководный (немногочисленный), южный типичный (многочисленный)) и в некоторых различиях в биологических характеристиках с этим связанных. Хотя можно признать это как начальный этап видообразования уже у рипуса.
Причиной же столь явного расхождения по морфологическим показателям между мелкими формами ряпушки Онежского и Ладожского озер, на наш взгляд, является то, что ряпушка представленных озер имеет несколько крупных популяций (стад) отличающихся по габитусу. Для Ладожского озера Дятлов (2002) говорит о том, что различные стада ряпушки данного водоема, при определенном навыке, можно различать даже визуально и, тем более что мы имели дело с различными водоемами. Тем более, как известно, ряпушка, являясь пластичным видом, может образовывать формы, резко отличающиеся других в зависимости от сложившихся условий существования.
Таким образом, ряпушка в зависимости от условий существования в водоеме, может образовывать формы, имеющие явные отличия от каких-либо других популяций и иметь признаки присущие только ей, как результат приспособления к сложившимся условиям существования.
Основная проблема видообразования состоит в том, чтобы объяснить происхождение разрывов между симпатрическими видами. Эти разрывы вполне ясно выражены и поддерживаются при помощи изолирующих механизмов. До тех пор пока виды считаются статичными, типологическими и одномерными, невозможно решить проблему перехода через эти разрывы. Особенностью симпатрического видообразования является то, что оно проводит к возникновению новых видов, всегда морфофизиологически близких к исходному виду или к, так называемым, видам-двойникам. Изолят не всегда происходит от популяций основателей. Иногда они образуются в результате распадения ранее непрерывного видового ареала на изолированные области. Действие мутаций, рекомбинаций и отбора в двух таких областях будет впредь различно и независимо, так что неизбежна все возрастающая генетическая дивергенция. Сколь быстро она будет происходить и приведет ли в результате к видообразованию – определяется в основном размером популяции. В большой популяции генетическое изменение, возможно, ведущее к достижению уровня вида, будет происходить очень медленно, если только видообразованию не благоприятствуют условия среды и генетическая структура изолированной популяции. Cтепень морфологической обособленности, достигнутая в период изоляции, не обязательно точно соответствует степени общей генетической дивергенции или, точнее говоря, репродуктивной изоляции. По мнению отдельных авторов (Абросов, 1987) наличие больших глубин в водоеме с незаполненными экологическими нишами может, сопровождается бурным видообразованием.
Любая резкая генетическая перестройка, подобная той, что может происходить в изолированных популяциях, особенно если они подвергаются явному экологическому сдвигу, может также одновременно затронуть генетическую основу изолирующих механизмов. Конечно же, интерпретировать экологическую изменчивость видов как механизм симпатрического видообразования нельзя. Она создает лишь предпосылки для адаптивных механизмов, которые необязательно приведут к каким-либо изменениям в фенотипе или генотипе. Однако она может облегчить и ускорить процесс видообразования.
Поводя итоги вышесказанному, можно заключить, что возможной основой симпатрического видообразования у рыб являются внутривидовые группировки, характеризующие различия в первую очередь биологических показателей (темп роста и.т.п.) в следствие, к примеру, увеличение мест обитания. Различия в размерах в дальнейшем, не исключено, определяет пищевую специализацию, а также неравномерность полового созревания. Из них первое явление снижает напряженность пищевых взаимоотношений, второе – затрудняет панмиксию и благоприятствует дальнейшей дивергенции. Подобный процесс в ряде поколений может привести к видовому обособлению внутривидовых форм, без какой бы то ни было географической изоляции. На основании результатов, дискриминантного анализа, по нашему мнению, следует пересмотреть систематический статус кильца и рипуса соответственно Онежского и Ладожского озер.
Библиографический список
- Абросов В. Н. О видообразовании в озерах. М., Наука 1987. 86 с.
- Бердников В.А. Основные факторы макроэволюции // Новосибирск: Наука Сиб. Отд-е. 1990. -253 с.
- Дятлов М.А. Рыбы Ладожского озера. - Петрозаводск. 2002. 280с.
- Майр Э. Популяция, виды и эволюция. М.: Мир, 1974. 460 с.
- Мина М.В. Микроэволюция рыб. Эволюционные аспекты фенетического разнообразия. М., Наука 1986. 205 с.
- Никольский Г.В. Структура вида и закономерности изменчивости рыб. М.: Пищевая промышленность, 1980. 335 с.
- Шмидт-Ниельсен К. Размеры животных: почему они так важны? - М.: Мир, 1987. - 259 с.
- Яблоков А.В., Юсуфов А.Г. Эволюционное учение. М.: Высшая школа, 1981. 343 с.
- Mayr E. Speciation and macroevolution // Evolution. -Vol. 36. 1982. P. 1119-1132.