

Известно, что болезни щитовидной железы (ЩЖ) являются одной из самых распространённых форм патологии человека. В настоящее время одними из наиболее распространённых патологий щитовидной железы являются её дисфункции – гипотиреоз и гипертиреоз (ГПРТ). ГПРТ (диффузный токсический зоб, базедова болезнь, тиреотоксикоз, болезнь Грэвса) является клиническим синдромом, обратным гипотиреозу, и характеризуется повышенной выработкой гормонов ЩЖ в организме. При ГПРТ полиморфные фолликулы ЩЖ приобретают неправильные контуры, а их просвет – щелевидную или неправильную форму в связи с тем, что выстилающий их тиреоидный эпителий, разрастаясь, образует утолщение фолликулярной стенки (так называемые сандерсоновские подушечки) или сосочки, нередко сильно разветвлённые, вдающиеся в просвет фолликулов (интрафолликулярная гиперплазия тиреоидного эпителия). Коллоид, содержащийся внутри фолликулов, в таких ЩЖ преимущественно жидкой консистенции и часто пронизан многочисленными различной величины резорбционными вакуолями. Клетки фолликулярного эпителия приобретают высокопризматическую или цилиндрическую форму (особенно в сандерсоновских подушечках), кровеносные сосуды резко полнокровны, строма обычно умеренно развита. В большом количестве встречаются интерфолликулярные островки или комплексы, образованные сочными клетками тиреоидного эпителия [1, 20]. Для ГПРТ характерно постоянное повышение температуры тела, частые головные боли, учащенное сердцебиение, повышение артериального давления. Также у больного может отмечаться эмоциональная неуравновешенность, мелкое дрожание рук, век, а также всего тела, склонность к поносам и повышенный аппетит. Эмоциональная неуравновешенность дает о себе знать в виде частой плаксивости, чрезмерной возбудимости, нарушений сна, а также быстрой сменой настроения [18, 21]. Интересен механизм образования базедова зоба, механизм увеличения ЩЖ при ГПРТ. Усиленное усвоение йода из пищи в кишечнике приводит к повышению уровня йода в крови. ЩЖ увеличивает продукцию тиреоидных гормонов, повышается их уровень в плазме крови. Гипофиз реагирует на это снижением секреции и выделения в кровь тиреотропного гормона (ТТГ). ЩЖ, сообразно с новым уровнем ТТГ в плазме крови, казалось бы, должна снизить секрецию тиреоидных гормонов. Но она этого сделать не может. Йод непрерывно в избытке поступает с кровью к ЩЖ, рецепторы ее клеток с максимальной возможностью захватывают излишний йод из крови и отправляют его в клетки ЩЖ для синтезирования гормонов. Их уровень в крови постоянно увеличивается. Гипофиз изо всех сил продолжает снижать уровень ТТГ для трго, чтобы вызвать снижение синтеза тиреоидных гормонов ЩЖ. Но теперь железа совсем не реагирует на команды гипофиза и на упавший ниже предельной нормы ТТГ в крови. Теперь ЩЖ переполнена йодом и стремится избавиться от его излишков. Синтез тиреоидных гормонов заметно усиливается, и увеличивается выделение их в кровь: по-другому избавляться от лишнего йода железа не умеет. Для усиленной продукции гормонов необходимо и дополнительное количество белка. Данный белок ЩЖ берёт из крови. Необходимо ещё и дополнительное количество коллоида – его продуцирует сама ЩЖ. В результате, ЩЖ от таких сверхнагрузок увеличивается в размерах.
У человека и других млекопитающих выделяют легкую, среднюю и тяжелую формы течения ГПРТ. При лёгкой форме похудание выражено умеренно, тахикардия не превышает 100 ударов в 1 мин., ритм сердечных сокращений не изменяется, признаков нарушения функций желез внутренней секреции (кроме ЩЖ) не отмечают. ГПРТ средней тяжести характеризуется выраженным похуданием, тахикардией, достигающей 100—120 ударов в 1 мин (особенностью тахикардии является ее стабильный характер, не зависящий от положения тела больного, предшествующего сна или длительного периода покоя), кратковременными изменениями сердечного ритма, нарушениями углеводного обмена, желудочно-кишечными расстройствами (частым жидким стулом), снижением концентрации холестерина в крови, постепенно нарастающими признаками надпочечниковой недостаточности. Тяжёлый ГПРТ является результатом длительно не леченного или плохо леченного ГПРТ. При этой форме отмечаются тяжелые нарушения функции отдельных органов и систем [1].
В коррегировании вышеотмеченных нарушений особого внимания заслуживает применение холиновых эфиров и амидов аминокислот и пептидов, заслуживающих существенного внимания с точки зрения особенностей их синтеза и биологической активности, играющих важную роль, благодаря их сходству по химической структуре к нейротрансмиттеру ацетилхолину, в системе нейроэндокринной регуляции клетки [5, 6]. Согласно результатам исследований последних лет [24, 25] холиновыми эфирами осуществляется ряд важнейших функций в растениях, а также в организме человека и животных. Вместе с тем продолжают отсутствовать сведения относительно применения эфиров холина при ГПРТ и результатов их действия на мотонейроны (МН) спинного мозга (СМ) у млекопитающих. Исходя из поиска оптимальных средств, стимулирующих и благоприятствующих лечению данных весьма сложных и распространённых патологий организма человека и животных и с учётом вышеотмеченных особенностей холиновых эфиров и амидов, нами проведён синтез трёх производных холина – йодметилата 2 – (диметиламино) этилового эфира N – (п – метоксибензоил) – DL – фенилаланина (ДЭФ), йодметилата 2 – (диметиламино) этилового эфира N – бензоил – DL – валина (ДЭВ) и йодметилата 2 – (диметиламино) этилового амида N – (п – метоксибензоил) – DL – фенилаланина (ДАФ) с последующим электрофизиологическим исследованием их действия на МН СМ у гипертиреоидных крыс. В данной работе подводятся итоги длительного этапа исследований реакции нейрональных элементов СМ крыс на сверхслабые воздействия синтетическими производными холина в условиях экспериментально вызванного ГПРТ.
Материал и методы. Эксперименты были поставлены на 100 крысах – самцах, массой 220 – 250 г, разделённых на следующие подопытные группы: первая – интактные животные – 10 экземпляров; вторая – животные с экспериментальным ГПРТ – 20 экземпляров; третья – животные с экспериментальным ГПРТ, получавшие внутримышечные инъекции ДЭФ в дозе 10-15 М – 20 экземпляров; четвёртая – животные с экспериментальным ГПРТ, получавшие внутримышечные инъекции ДЭВ в дозе 10-15 М – 20 экземпляров; пятая – животные с экспериментальным ГПРТ, получавшие внутримышечные инъекции ДАФ в дозе 10-15 М – 30 экземпляров. Исследуемые соединения вводились подопытным животным в течение 1 месяца. ГПРТ вызывался каждодневным систематическим введением подопытным животным гормона ЩЖ L – тироксина в дозе 500 мкг/кг массы тела в течение 18 дней. За каждым животным в период дачи препаратов велось индивидуальное наблюдение с каждодневной регистрацией клинических показателей. Последующие электрофизиологические исследования биоэлектрических показателей СМ проводились в остром эксперименте. В острых опытах под эфирным наркозом крысу обездвиживали дитилином и переводили на искусственное дыхание; СМ пересекался под новокаином ультразвуковым ножом на уровне Т2 – Т3. После прочной фиксации пояснично-крестцового отдела позвоночника в стереотаксическом приборе производили ламинэктомию данной области. Электрофизиологическими методами изучалась внеклеточная фоновая (ФА) и вызванная (ВА) электрическая активность одиночных МН СМ у интактных и гипертиреоидных животных. Регистрацию и анализ внеклеточной ФА и ВА одиночных МН СМ производили с помощью специальных программ на компьютере. При записи ВА одиночных МН СМ проводилась оценка стационарности престимульной и постстимульной активности нейрона после электрического раздражения седалищного нерва. Выборка спайков проводилась с помощью амплитудного дискриминатора посредством программного анализа. Вычислялась скользящая частота (для каждых 10 межимпульсных интервалов с шагом 5 интервалов), сериальные гистограммы до 50 порядка, гистограммы межимпульсных интервалов на основе которых вычислялась средняя частота фонового нейронального потока одиночных клеток, а также спайковая выборка нескольких (до 20) повторений до и после раздражения. Последующий анализ ФА и ВА одиночных МН СМ крыс осуществляли по алгоритму, подробно описанному в наших предыдущих статьях [2, 4, 7 – 9, 10].
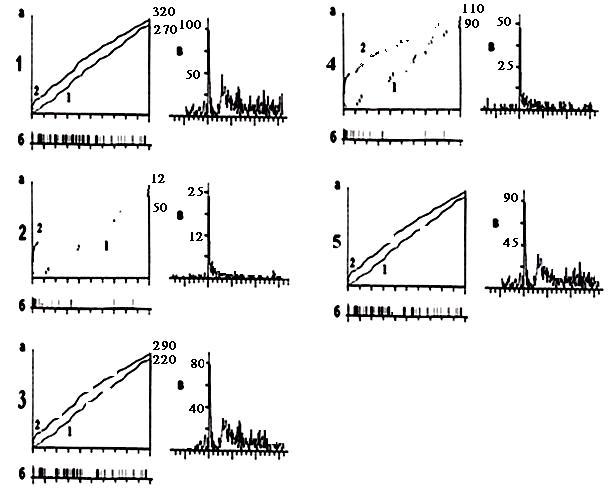
Рис.1. Примеры кумулятивных (а) и суммированных (в) пре – и постстимульных гистограмм внеклеточной фоновой и вызванной активности одиночных мотонейронов вентрального рога спинного мозга крыс в норме (1 а, б, в); у гипертиреоидных крыс (2. а, б, в); у гипертиреоидных крыс, получавших ДЭФ (3, а, б, в); у гипертиреоидных крыс, получавших ДЭВ (4 а, б, в); у гипертиреоидных крыс, получавших ДАФ (5 а, б, в). Глубина отведения всех 5 мотонейронов – 1400 микрон. На «а»: ордината – число импульсов до и после стимуляции нерва, абсцисса – время регистрации импульсного потока. На «б»: картина импульсного потока после стимуляции нерва в избранном интервале времени. На «в»: ордината – процент импульсов (в бинах) от числа проб, абсцисса – последовательность бинов.
Результаты и обсуждение. На рисунке 1 приведены примеры кумулятивных (рисунок 1, пункты 1 – 3, а, б) и суммированных (рисунок 1, пункты 1 – 3, в) престимульных и постстимульных гистограмм внеклеточной ФА и ВА одиночного МН СМ у интактных животных (глубина 1400 микрон; пункт 1, а, б, в); у гипертиреоидных животных (глубина 1400 микрон, пункт 2, а, б, в); у гипертиреоидных животных, получавших ДЭФ (глубина 1400 микрон, пункт 3, а, б, в); у гипертиреоидных животных, получавших ДЭВ (глубина 1400 микрон, пункт 4, а, б, в); у гипертиреоидных животных, получавших ДАФ (глубина 1400 микрон; пункт 5, а, б, в) в указанных выше дозировках. При экстраклеточной регистрации ФА и ВА одиночных МН СМ у гипертиреоидных крыс наблюдается трансформация регулярного типа разряда ФА одиночного МН в пачечную «патологическую» активность (рисунок 1, пункт 2, а, кривая 1) с почти полным исчезновением вызванного синаптического ответа (рисунок 1, пункт 2, а, кривая 2), вызванное, вероятно, патологическим влиянием ГПРТ. После введения ДЭФ наблюдается эффект стабилизации пачечной активности одиночного МН СМ с постепенным переходом в нормальный регулярный тип разряда МН с сохранением и увеличением вызванного синаптического ответа (рисунок 1, пункт 3, а, кривые 1, 2), что говорит об усилении действия ДЭФ. Что же касается импульсного потока (рисунок 1, пункт 3, б), то здесь, после введения ДЭФ (доза 200 мкг/кг массы тела), он представлен регулярным видом фонового нейронального разряда МН. Аналогичные сдвиги вызванного ответа при ГПРТ в 2 – 3 раза отражены и на суммарных гистограммах (рисунок 1, пункт 2, в), а при действии ДЭФ – на суммарных гистограммах (рисунок 1, пункт 3, в). После введения ДЭВ в дозе аналогичной ДЭФ (рисунок 1, пункт 4, а, кривые 1, 2) наблюдается частичная трансформация пачечного типа ФА и ВА одиночного МН СМ в регулярный тип разряда; в картине импульсного потока (рисунок 1, пункт 4, б) также продемонстрирован регулярный вид фонового разряда МН, но, по – сравнению с картиной импульсного потока, полученной при применении ДЭФ (рисунок 1, пункт 3, б), восстановление фонового нейронального разряда одиночного МН СМ выражено гораздо слабее, что позволяет заключить о более слабо выраженном действии ДЭВ на фоновую ритмику одиночных МН СМ крыс по сравнению с ДЭФ. На суммарных гистограммах (рисунок 1, пункт 4, в) при применении ДЭВ отражены сдвиги вызванного ответа одиночного МН СМ в 1 – 2 раза, что в лишний раз подтверждает более слабое протекторное действие ДЭВ по сравнению с ДЭФ. После введения ДАФ в дозе аналогичной ДЭФ и ДЭВ (рисунок 1, пункт 5, а, кривые 1, 2) наблюдается более усиленный эффект трансформации пачечного типа ФА и ВА одиночного МН СМ по сравнению с ДЭФ и ДЭВ; в картине импульсного потока (рисунок 1, пункт 5, б) показан эффект восстановления практически до нормы фонового нейронального разряда с переходом его в регулярный тип ответа МН. На суммарных гистограммах при применении ДАФ представлено резкое усиление вызванного ответа одиночного МН СМ в 5 – 6 раз (рисунок 1, пункт 5, в), что говорит о более сильном, активирующем действии ДАФ на вызванную ритмику одиночных МН СМ по сравнению с ДЭФ и ДЭВ. Анализируя данный рисунок можно сделать вывод об активирующем, протекторном действии сверхмалых доз (СМД) холиновых производных на внеклеточную ФА и ВА одиночных МН СМ крыс при экспериментальной дисфункции ЩЖ – ГПРТ.
Подводя итоги данного исследования можно, считаем целесообразным отметить, что при изучении действия СМД холиновых производных при экспериментальной патологии ЩЖ крыс – ГПРТ, нами был получен весьма неожиданный результат. Первоначальная доза каждого исследуемого препарата (10-3М) была не только активной для МН, но и довольно токсичной, поэтому пришлось перейти на менее концентрированный раствор. Доза на четыре порядка ниже первоначальной оказалась не только менее токсичной, но и более эффективной. Дальнейшее уменьшение концентрации привело к росту эффекта, он достигал максимума (при 10-15 М), затем снижался до уровня (при 10-17 М), практически совпадающего с контрольными результатами. Исходя из вышеизложенного, оптимальной дозой препаратов, использующихся для лечения ГПРТ, мы считаем дозу 10-15 М. Уровень биологической организации, на котором проявляется действие СМД биологически активных веществ (БАВ), также весьма разнообразен – от макромолекул, клеток, органов и тканей до животных, растительных организмов и даже популяций. Сказанное не означает, что эффект наблюдался при СМД любого БАВ на любом биологическом объекте. Следует отметить, что получение эффекта при действии вещества в концентрациях 10-13 – 10-17 М и ниже нельзя, вероятно, связать с какой – то определённой структурой вещества или ступенью биологической организации. Из литературных данных [12 – 13, 16] и из наших собственных исследований можно сделать вывод, что в проявлениях влияния на клеточный метаболизм СМД БАВ обнаруживают много общих особенностей, которые касаются как формальных признаков (дозовые зависимости), так и показателей биологической активности. Природа этого феномена может быть связана с общностью критических мишеней, например, клеточных и субклеточных мембран, а также с особенностями кинетики реакций, в которых важную роль играют слабые взаимодействия. К числу характерных для эффектов СМД свойств следует отнести:
- немонотонную, полимодальную зависимость «доза – эффект». В большинстве случаев максимумы активности наблюдаются в определённых интервалах доз, разделённых между собой так называемой «мёртвой зоной»;
- изменение чувствительности (как правило, увеличение) биообъекта к действию разнообразных агентов как эндогенных, так и экзогенных (последние могут быть как той же, что в случае воздействия СМД, так и иной природы);
- проявление кинетических парадоксов, а именно возможность уловить эффект СМД БАВ, когда в клетке или организме имеется то же вещество в дозах на несколько порядков выше, а также влияние на рецептор вещества в дозах на порядки более низких, чем константы диссоциации комплекса лиганд – рецептор;
- зависимость «знака» эффекта от начальных характеристик объекта;
- «расслоение» свойств БАВ по мере уменьшения его концентраций, при котором ещё сохраняется активность, или исчезают побочные эффекты.
Общие закономерности влияния сверхмалых доз препаратов наиболее ярко проявляются при изучении дозовых зависимостей. В некоторых случаях эта зависимость бимодальная: эффект возрастает при СМД препаратов, затем при увеличении дозы уменьшается, сменяется «мёртвой зоной» и вновь усиливается. Иногда в дозовой зависимости обнаруживается стадия «перемены знака» эффекта. Например, если в области СМД отмечалась ингибирующая активность, то по мере роста концентрации она сменялась на стимулирующую, а затем вновь проявлялся ингибирующий эффект [3, 23].
В результате проведенного анализа полученных данных мы пришли к выводу, что в целом имеется положительный эффект от применения СМД холиновых производных при ГПРТ, в особенности дозы 10-15 М. Итак, результаты ранее проведенных исследований по изучению действия холиновых производных [11, 14 – 15, 19, 22, 26], а также результаты данной серии исследований показали эффективность действия СМД холиновых производных, в особенности ДАФ, на внеклеточную ФА и ВА одиночных МН СМ крыс при экспериментально вызванном ГПРТ.
Библиографический список
- Бреславский А. С., Гордиенко В. М. Патологическая анатомия желёз внутренней секреции. Изд. «Здоров’я», Киев, 1974, с. 34 – 35.
- Киприян Т. К., Хачатрян Т. С. Протекция гипоталамическим нейрогормоном острой нейродегенерации мотонейронов спинного мозга, вызванной змеиными ядами. Ж. Информационные технологии и управление, 2007, № 5, с. 68 – 75.
- Матинян Л. А., Нагапетян Х. О., Андреасян А. С., Киприян Т. К., Хачатрян Т. С. Об усилении некоторых целебных воздействий. Ж. Вестник МАНЭБ, 2007, т. 12, № 4, вып. 2, с. 157 – 159.
- Матинян Л. А., Хачатрян Т. С., Киприян Т. К., Арутюнян Э. Ю., Авакян А. Э. Влияние трипсина, химотрипсина, лидазы на вызванную активность повреждённых травмой одиночных мотонейронов спинного мозга крыс. Ж. Вестник МАНЭБ, 2008, т. 13, № 4, вып. 1., с. 117 – 120.
- Мнджоян О. Л., Топузян В. О. Методы синтеза и свойства β – диметиламиноэтиловых и холиновых эфиров аминокислот и пептидов. Ж. Успехи химии., L (12): 2198 – 2211. 1981.
- Ткачук В. А. Молекулярные механизмы нейроэндокринной регуляции. Ж. Соросовский образовательный журнал, 1998, № 6, c. 25 – 31.
- Хачатрян Т. С. Действие лидазы и тироксина на фоновую электрическую активность одиночных пирамидных нейронов коры больших полушарий крыс. Ж. Биолог. журн. Армении, 3 – 4 (59), 2007, с. 198 – 202.
- Хачатрян Т. С. Протекция тироксином изменений вызванной активности повреждённых травмой одиночных мотонейронов спинного мозга крыс. Ж. Биолог. журн. Армении, 2008, № 3,т. LX, с. 64 – 67.
- Хачатрян Т. С., Киприян Т. К. Изменение суммарной и одиночной электрической активности интернейронов и мотонейронов спинного мозга крыс при гипотиреозе и гипертиреозе. Ж. Информационные технологии и управление, 2006, № 4 – 3, с. 46 – 51.
- Хачатрян Т. С., Нагапетян Х. О., Матинян Л. А. Фитотерапия в сочетании с тироксином при органических повреждениях спинного мозга. Ж. Биолог. журн. Армении, 2008, № 4,т. LX, с. 51 – 55.
- Brown M., Davies I. M., Moffat C. F., Redshaw J., Craft J. A. Characterisation of choline esterases and their tissue and subcellular distribution in mussel (Mytilus edulis). J. Mar. Environ. Res., 2004, № 57, v. 3, pp. 155 – 169.
- Degliesposti A. Choline hydrochloride therapy of hyperthyroidism. J. Policlinico Part., 1952, № 59, v. 25, pp. 839 – 841.
- Degliesposti A. Effects of choline in hyperfunctioning thyroid adenoma in a child. J. Policlinico Part., 1952, № 59, v. 25, pp. 837 – 839.
- Di Venosa G., Hermida L., Battle A., Fukuda H., Defain M. V., Mamone L., Rodriguez L., MacRobert A., Casas A. Characterisation of liposomes containing aminolevulinic acid and derived esters. J. Photochem. Photobiol., 2008, № 92, v. 1, pp. 1 – 9.
- Eibl K. H., Lewis G. P., Betts K., Linberg K. A., Gandorfer A., Kampik A., Fisher S. K. The effect of alkylphosphocholines on intraretinal proliferation initiated by experimental retinal detachment. J. Invest. Ophtalmol. Vis. Sci., 2007, № 48, v. 3, pp. 1305 – 1311.
- Giardino L., Giuliani A., Bettelli C., Calza L. Thyroid hormone and retinoids affect motoneuron phenothype and reaction after axothomy in the spinal cord of adult rats. J. brain Res., 2002, № 925, v. 2, pp. 122 – 132.
- Hamplova B., Novakova O., Tvrzicka E., Pelouch V., Novak F. Effect of hypo – and hyperthyroid states on phospholipid composition in developing rat heart. J. Mol. Chem. Bioche., 2003, № 252, v. 1 – 2, pp. 295 – 303.
- Holmes – McNary M. Q., Cheng W. L., Mar M. H., Fussel S., Zeisel S. H. Choline and choline esters in human and rat milk and in infant formulas. J. Am. J. Clin. Nutr., 1996, № 64, v. 4, pp. 572 – 576.
- Jonderko G., Straszecka J., Marcisz C., Wieczorek U. Influence of treating hypothyroidism and hyperthyroidism upon physical reaction time. J. Pol. Arch. Med., 1992, v. 5, № 88, pp. 295 – 301.
- Jorde R. “Subclinical” thyroid disease. J. Tidsskr. Nor. Lageforen., 2002, № 122, v. 9, pp. 938 – 940.
- Masson P., Froment M. T., Gillon E., Nachon F., Lockridge O., Schopfer L. M. Hydrolysis of oxo- and thio-esters by human butyrylcholinesterase. J. Biochim. Biophys. Acta, 2007, № 1774, v. 1, pp. 16 – 34.
- Mellon E. A., Beesam R. S., Kasam M., Baumgardner J. E., Borthakur A., Witschey W. R. Jr., Reddy R. Single shot T1rho magnetic resonance imaging of metabolically generated water in vivo. J. Adv. Exp. Med. Biol., 2009, № 645, pp. 279 – 286.
- Terkeltaub R. Physiologic and pathologic functions of the NPP nucleotide pyrophosphatase/phosphodiesterase family focusing on NPP1 in calcification. J. Purinergic Signal., 2006, № 2, v. 2, pp. 371 – 377.
- Walz I., Schwack W. Multienzyme inhibition assay for residue analysis of insecticidal organophosphates and carbamates. J. Argic. Food. Chem., 2007, № 55, v. 26, pp. 10563 – 10571.
- Zeisel S. H. Choline: Needed for Normal Development of Memory. J. Am. J. Clin. Nutr., 2000, № 905, v. 19, pp. 528 – 531.
Количество просмотров публикации: Please wait